
Small molecules come in all sorts of different shapes and are capable of wonderful things. Of particular significance are those privileged types of small molecules that interact with their protein targets with high ligand efficiency. I will dedicate a future blog post to the concept of ligand efficiency, but I am sure that we all “feel” what it means without any formal definition. Ligand efficiency is a measure of how well a given molecule engages its innate features when it interacts with a protein target. Understanding the balance of enthalpy and entropy in this type of molecular recognition is paramount in probe/drug design. Great strides have been made in explaining (through docking simulations) how and why organic molecules interact with their targets. A lot of the developed algorithms work extremely well (Schrodinger’s Glide package is my favorite at the moment). The simulations may not have the best predictive power, but they are certainly capable of explaining experimental facts. Or are they?
I give you one of the workhorses of molecular biology: biotin/streptavidin interaction. Everyone is aware of the significance of this non-covalent “glue”. There are many examples of affinity experiments that utilize the strength of biotin/streptavidin pair. Personally, I have been in awe of this system because I just don’t get it: how the heck is such tight binding possible? You look at the dinky little biotin molecule and there is just no way to expect that its binding interaction with streptavidin would be on the order of 10-14 mol/L (I mean the dissociation constant, Kd). It turns out that sophisticated modern simulations have a heck of a trouble predicting this extreme ligand efficiency. The experimentally determined binding affinity is ORDERS of magnitude higher than predicted. So what is the basis for this impressive affinity?
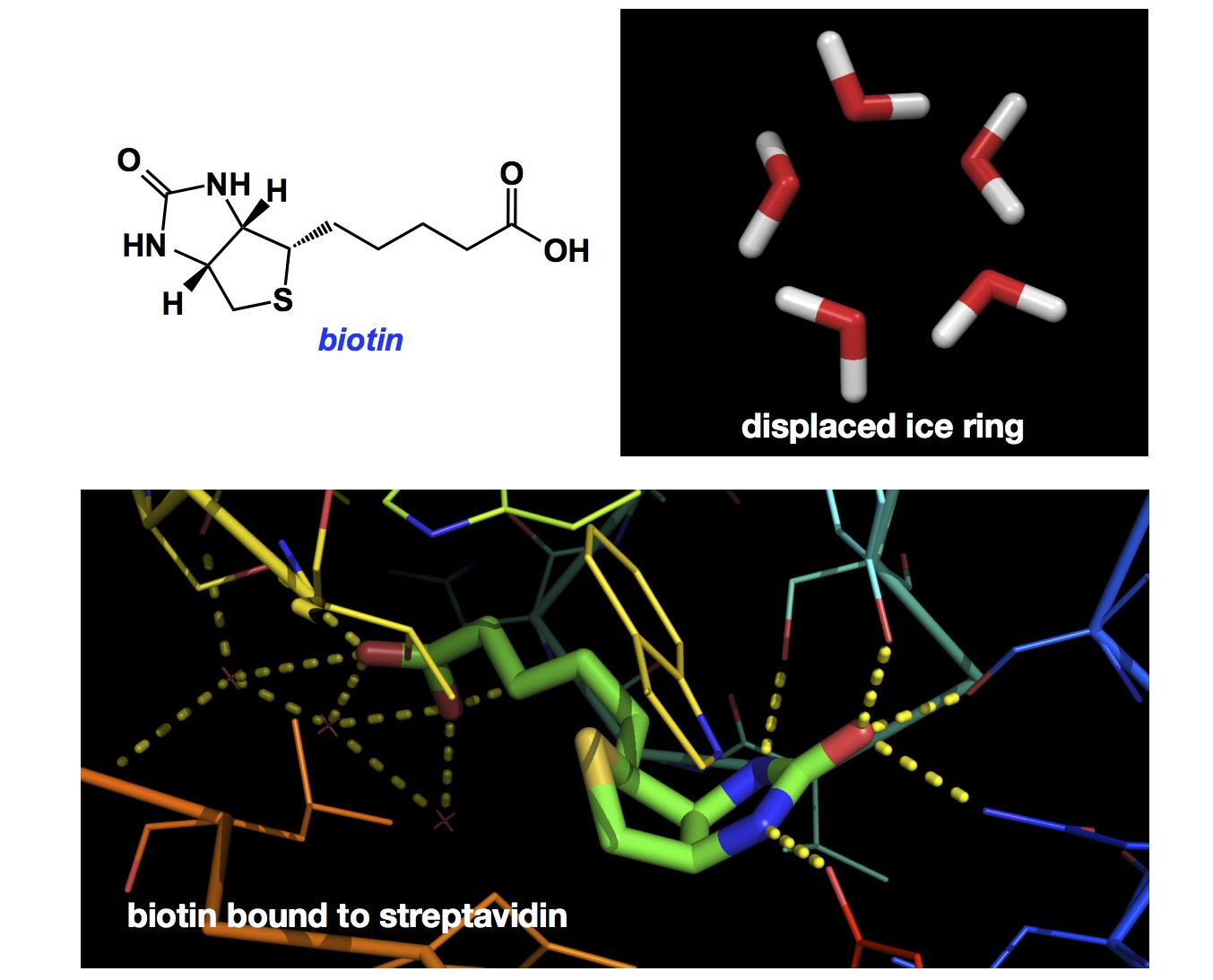
It turns out that the reason likely lies in the amazing five-membered, ice-like ring of water molecules that is present in the binding site of streptavidin. Biotin is uniquely geared towards displacement of this “ice-ring” arrangement from the binding site, which generates a huge entropic driving force (see the graphic above showing the ring and the ultimate structure where biotin is in its place). There is still no way to predict this kind of behaviour computationally, I am afraid. The lesson here is that displacement of ordered waters is a VERY attractive way towards designing super-ligands. Berne and co-workers have published a very insightful PNAS paper on this subject several years ago (I thank Robert Campbell of Queens University for bringing this work to my attention):
