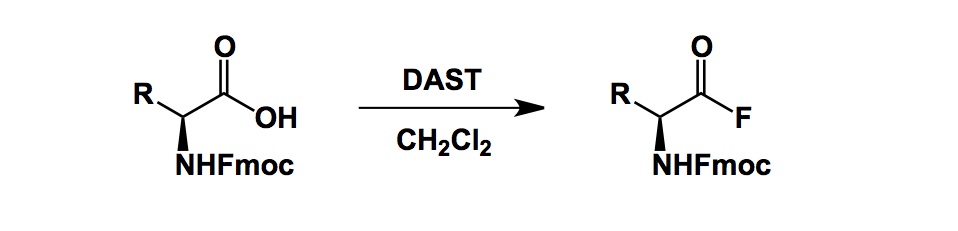
The challenge of oxidative nitrogen transfer is fascinating because nature does not know how to do it, at least not when it comes to oxidative C-N bond formation. I already commented on this feature of biosynthesis in the past and there is no need to belabor the point. The arsenal of synthetic tricks we have is astounding. The trouble is, of course, that generation of nitrogen oxidants can require rough conditions that might adversely affect other “spinach” that hangs off your molecule. We worked on this problem in the past and found electrosynthesis to be full of options. Here is an old pic of one of our reactions along with the reactor we used. I am only indicating the process taking place at the anodic compartment, in which we oxidized the hydrazine-containing starting material. The cathodic compartment separated by a frit you see, is colorless. It is actually possible to see hydrogen bubbles on this picture – they come as a result of proton reduction (protons are produced during oxidation). I am also attaching a link to the JACS paper where we demonstrated how anodic electrochemistry helped us solve some challenging problems of oxidative nitrogen transfer. 
http://pubs.acs.org/doi/pdf/10.1021/ja0172215
When I visited the Dallas ACS meeting a couple of weeks ago, I heard Phil Baran speak about his lab’s use of electrochemistry in natural product total synthesis. I saw his JACS communication earlier today and enjoyed reading this work because it brought back many fond memories of hydrazine chemistry. As was the case in our oxidative approach to aziridines, the beauty of electrochemistry in Phil’s case is in offering a glimpse of synthetic opportunity by judiciously choosing the right window of applied anodic potential at which the amine can be selectively oxidized. In this particular case, electrooxidation leads to the formation of a hydrazine-containing natural product dixiamycin B. Any chemical oxidants Phil’s lab tried, failed in this oxidation reaction. I was not surprised that carbon was the best electrode material in this case. It is almost certain that the “flat-ish” substrate you see interacts quite well with the anode surface. We could not use carbon in our aziridine chemistry, though. Platinum was the key to us as graphite led to strong background currents corresponding to alkene oxidation.